リブロース1,5-ビスリン酸カルボキシラーゼ/オキシゲナーゼ
リブロース-1,5-ビスリン酸カルボキシラーゼ/オキシゲナーゼ (ribulose-1,5-bisphosphate carboxylase/oxygenase) はカルビン - ベンソン回路において炭酸固定反応に関与する唯一の酵素である(EC番号は 4.1.1.39)。リブロース1,5-ビスリン酸に二酸化炭素を固定し2分子の3-ホスホグリセリン酸を生成する反応を触媒する。植物に大量に含まれ、地球上で最も多いタンパク質ともいわれる。具体的にはホウレンソウの葉の可溶性タンパク質の5-10%は本酵素に占められる。

リブロース-1,5-ビスリン酸カルボキシラーゼ/オキシゲナーゼ本来の生理学的な役割はリブロース 1,5-ビスリン酸 (RuBP) へのカルボキシル化(カルボキシラーゼ反応)であるために、リブロース-1,5-ビスリン酸カルボキシラーゼが正しい呼称である。しかし本酵素は植物の炭素固定反応を律速している主原因となるリブロース 1,5-ビスリン酸へのオキシゲナーゼ作用(オキシゲナーゼ反応)が特徴的であり、この両反応の競合関係にあるためリブロース-1,5-ビスリン酸カルボキシラーゼ/オキシゲナーゼと呼称されることが多い。
呼称の長さから Ribulose 1,5-bisphosphate carboxylase/oxygenase の各文字をとって RubisCO と表記されることが多い。他の別名として、リブロース1,5-ビスリン酸カルボキシラーゼ、RuBPカルボキシラーゼ、Rubisco、RuBisCO、ルビスコなど。
カルボキシラーゼ反応
編集RubisCOは生理学的に以下の反応を触媒する。括弧内は炭素数を意味する。
- D-リブロース1,5-ビスリン酸 (C:5) + CO2 (C:1) + H2O → 2 × ホスホグリセリン酸 (C:3 × 2)
この反応の補因子としてマグネシウムイオン (Mg2+) を要求する。この反応の平衡は著しく右方向に寄っており、逆反応はほとんど起きない。この反応はカルビン - ベンソン回路におけるカルボキシル化過程であり、本代謝系に唯一固有な反応である。カルビン-ベンソン回路における他の反応は、すべて解糖系あるいはペントースリン酸経路に見られる。したがってカルビン-ベンソン回路はRubisCOのカルボキシラーゼ反応によって調節される。RubisCOによるカルビン-ベンソン回路の調節については後述する。

カルボキシラーゼ反応は詳細に分類すると以下のステップを経る。
- D-リブロース1,5-ビスリン酸がRubisCOの作用によりエンジオール型中間体となる。
- RubisCOにCO2が結合する(この二酸化炭素は基質ではない)。
- RubisCO-CO2複合体にMg2+が結合し活性型となる。
- エンジオール型中間体のリブロース1,5-ビスリン酸にCO2が付加し(この二酸化炭素が基質となる)、3-オキソ中間体となる。
- 3-オキソ中間体のリブロース1,5-ビスリン酸が加水分解され2分子のホスホグリセリン酸を生じる。
- 2分子のホスホグリセリン酸のうち1分子のみがCO2由来の炭素原子を有する。
RubisCOはきわめて分子活性 (kcat) の低い酵素であり、3 s−1程度である(酵素1分子で1秒当たり3分子のCO2を固定する)。
カルビン-ベンソン回路の調節
編集暗所に置いた植物細胞に光照射すると炭酸固定速度がしばらく低い状態が続くが、次第に定常速度に到達する。これは、暗所でカルビン-ベンソン回路に関与する複数の酵素の活性が低下しているからである。RubisCOもそうした酵素のひとつである。しかしRubisCOは光そのものによって活性化されるわけではなく、以下の要素によって活性化される。
- Mg2+の存在
- pH 8.5程度の弱アルカリ
- 重炭酸イオン (HCO3−) の存在(ホモトロフィック効果)
- 2-カルボキシアラビニトール一リン酸量の低下
- ATP要求性RubisCO活性化酵素による活性化
光によるRubisCO活性化と深く関わっているのは弱アルカリによる活性化である。光照射により葉緑体において光化学反応が起き、チラコイド内にプロトン (H+) が取り込まれるが、このときストロマ内のpHが上昇する。RubisCOはストロマ内に局在しており、活性化を受ける。また、チラコイドのプロトンの取り込みと同時に膜電位上昇に伴うMg2+の輸送がチラコイドからストロマ側に行われ、ここでもRubisCOの活性化を促進する。チラコイド、ストロマなどの構造は葉緑体の項を参照。
RubisCOは基質である重炭酸イオンの濃度上昇によって最大速度が上昇し、本酵素がアロステリック酵素であることを示している。また、RubisCOのカルボキシラーゼ反応の阻害剤となりうる2-カルボキシアラビニトール一リン酸は夜間に存在量が多く、昼間には低下する。さらにATP要求性RubisCO活性化酵素も、光化学反応によりストロマ側のATP濃度が上昇が活性化の鍵となる。すなわち、暗所では全ての要素が逆転し、RubisCOのカルボキシラーゼ反応を阻害する方向に働く。
オキシゲナーゼ反応
編集強光下で光合成を行っていた植物細胞を21%O2濃度条件(通常の大気)における暗所に移動した場合、直後に大量のCO2放出が見られる。同様の実験を1.5%O2濃度条件の暗所に移動した場合、CO2の放出は観察されなくなる。明条件におけるCO2の放出を光呼吸というが、この原因になっている主反応がRubisCOのオキシゲナーゼ反応である。光呼吸は同化したCO2を再放出する過程であり(植物の生育速度の低下)、多くの主要作物(小麦、米、豆類など)でこの現象が見られることから収量上昇など経済的にも注目されている。RubisCOのオキシゲナーゼ反応は以下のとおりである。
- リブロース1,5-ビスリン酸 + O2 → 3-ホスホグリコール酸 + 3-ホスホグリセリン酸
一見、CO2は放出されていないように見えるが、3-ホスホグリコール酸がペルオキシソームおよびミトコンドリアを経て代謝されCO2を放出する。3-ホスホグリセリン酸はそのままカルビン-ベンソン回路にて還元過程に入る。また、実際にCO2を放出する反応のみならず、オキシゲナーゼ反応がカルボキシラーゼ反応と競合し阻害することによって、見かけのCO2放出量を増加させ、光合成能率の低下を招く。

オキシゲナーゼ反応は詳細に分類すると以下のステップを経る。
- D-リブロース1,5-ビスリン酸がRubisCOの作用によりエンジオール型中間体となる。
- RubisCOにO2が結合する。
- RubisCO-O2複合体にMg2+が結合し活性型となる。
- エンジオール型中間体のリブロース1,5-ビスリン酸にO2が付加し、3-オキソ型中間体となる。
- 3-オキソ型中間体リブロース1,5-ビスリン酸と水酸化物イオン (OH−) が反応し、3-ホスホグリコール酸、3-ホスホグリセリン酸および水酸化物イオンをそれぞれ1分子ずつ生じる。
- リブロース1,5-ビスリン酸に結合した酸素原子は3-ホスホグリコール酸の2位および放出される水酸化イオンに結合している。
上記のカルボキシラーゼ反応と比較して、反応ステップは極めて似通っており両反応が競合するのも、こうした酵素的な諸反応が類似していることが原因である。
RubisCOがオキシゲナーゼ活性を有する原因として、活性中心へのO2分子の取り込まれやすさが考えられる。RubisCOを有する光合成生物が出現した当初、地球上の大気組成は現在よりも還元的でCO2濃度が高かったと考えられており、こうした欠点は表在化しなかった。しかしながら、陸上植物の台頭により大気中の酸素分圧の上昇そして二酸化炭素分圧の低下を招き、RubisCOの欠点が光呼吸という形で現れてきたと考えられている。
RubisCOのカルボキシラーゼ/オキシゲナーゼ活性比とCO2/O2比の関係を表す値を「CO2補償濃度」といい、自然に起きている暗呼吸と光呼吸がつりあい、みかけのCO2放出速度が0になる。一般的なC3植物のCO2補償濃度は50–100 ppmである。また、CO2補償濃度以外のRubisCOの能力を示す値として、任意のCO2/O2濃度におけるカルボキシラーゼ/オキシゲナーゼ活性比である「τ値」が挙げられる。一般的に、陸上植物型のRubisCO (Form I) は嫌気性光合成細菌のRubisCO (Form II) に比べて、高酸素分圧条件に適応した痕跡が見られ、CO2補償濃度およびτ値において優れた値を示す。Form I、IIについては後述する。
RubisCOの生物界における分布
編集RubisCOを有している生物として陸上高等植物、藻類、渦鞭毛藻類など真核生物のほか、シアノバクテリア、嫌気性光合成細菌、化学合成独立栄養細菌といった多くの原核生物(細菌、古細菌)があげられる。RubisCOは一般的にForm IおよびForm IIに分類されるが、古細菌から見つかっているRubisCOはどちらのFormにも属さない第3のRubisCOを有する事が分かっている。また、最近RubisCOに必要なアミノ残基を欠くRubisCO-like proteinが細菌および古細菌の両方から見つかっている。
-
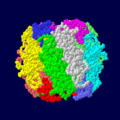 ホウレンソウRubisCOの空間充填モデル。Form I RubisCOは大サブユニットと小サブユニットからなるヘテロダイマーの8量体で構成される。
ホウレンソウRubisCOの空間充填モデル。Form I RubisCOは大サブユニットと小サブユニットからなるヘテロダイマーの8量体で構成される。 -
 光合成細菌 Rhodospirillum rubrum RubisCOのリボンモデル。Form II RubisCOはForm I RubisCOの大サブユニットに類似したサブユニットのホモ二量体から構成される。
光合成細菌 Rhodospirillum rubrum RubisCOのリボンモデル。Form II RubisCOはForm I RubisCOの大サブユニットに類似したサブユニットのホモ二量体から構成される。 -
 好熱性古細菌 Thermococcus kodakaraensis RubisCOのリボンモデル。Form III RubisCOはホモ10量体から構成される。
好熱性古細菌 Thermococcus kodakaraensis RubisCOのリボンモデル。Form III RubisCOはホモ10量体から構成される。 -
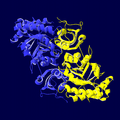 光合成細菌 Chlorobium tepidum RubisCO様タンパク質のリボンモデル。Form IVはRubisCOに必要ないくつかのアミノ酸を欠いている。
光合成細菌 Chlorobium tepidum RubisCO様タンパク質のリボンモデル。Form IVはRubisCOに必要ないくつかのアミノ酸を欠いている。




Form I
編集Form Iの特徴は以下のとおりである。
- 大サブユニット8個および小サブユニット8個からなるヘテロ16量体(全体の分子量は約550 kDa)。
- 大サブユニットの分子量は50–55 kDa、小サブユニットの分子量は12–18 kDa。
- 陸上高等植物、藻類、シアノバクテリアといった酸素発生型の生物が主に有している(しかしながら、嫌気性の化学合成独立栄養細菌からも多く見つかっている)。
- 陸上高等植物や緑藻類を含む緑色植物では、大サブユニット遺伝子 (rbcL) が葉緑体に、小サブユニット (rbcS) が核にそれぞれコードされている。それ以外の真核藻類、紅藻や不等毛植物、クリプト藻、ハプト藻等では両サブユニットとも葉緑体にコードされており、転写単位(オペロン)を形成している。
- CO2補償濃度、τ値においてより高酸素分圧下での活性に適応している。
Form II
編集Form IIの特徴は以下のとおりである。
- Form Iの大サブユニットに該当するサブユニットのホモ2量体からなる(全体の分子量は約100 kDa)
- 渦鞭毛藻類や紅色非硫黄細菌等が主として有している。渦鞭毛藻では核コードである。
- CO2補償濃度、τ値はForm Iに比べて高酸素分圧に適応していない。
- Form Iよりも古い系統に属すると考えられている。
Form III
編集古細菌型RubisCO (Form III) の特徴は以下のとおりである。
- 大サブユニットに該当すると考えられるサブユニットのホモ10量体である(ユーリ古細菌門に属する Thermococcus kodakaraensis のケース、分子量は500 kDa程度)。
- サブユニットのアミノ酸配列相同性が他のForm IおよびIIの大サブユニットと比較して著しく低い(Form Iと41%、Form IIと34%;ユーリ古細菌門に属するMethanocaldococcus jannaschii のケース)。
- Pyrococcus horikoshii、Archaeoglobus fulgidus などユーリ古細菌門から主として見つかっている。
Form IV
編集RubisCO-like protein (Form IV) の特徴は以下の通りである
- Chrolobium limicola、C. tepidum、A. fulgidus そして Bacillus subtilis と古細菌および細菌の両方から見つかっている。
- RubisCO触媒部位に必須とされる幾つかのアミノ酸を欠いている。
- C. tepidum にてノックアウト実験が行なわれたが硫黄代謝に欠陥が現れ、新規代謝系に関与していることが示唆されている。
立体構造
編集上述したようにRubisCOは地球上で最も存在量の多いタンパク質である。そのため、歴史的にも構造生物学的研究が進んだ酵素の一つであった。1971年に最初のRubisCO結晶が得られたものの、最初のX線結晶構造が明らかになったのは1990年のことであった。
まずはじめに得られた立体構造は紅色非硫黄細菌 Rhosdospirillum ruburm のForm II RubisCOである。解析の結果、Form IIは同一のサブユニット2個が対称性を持って結合している、分子量100 kDa程度のホモ2量体であることがわかった。Form I RubisCOの立体構造は同年にタバコ葉のものが明らかになった。Form Iは大サブユニット2個、小サブユニット2個のL2S2単位が計4個対称性を持って結合している分子量550 kDa程度のL8S8型であることが明らかになった。結晶化から立体構造の把握まで非常に時間がかかったことに関して、分子量が極めて大きく高い解像度を得られなかったことが原因と考えられている。
その後、多くの生物からForm I、II RubisCOの立体構造が明らかになった。また、基質や生産物を加えながら結晶化を行なうことにより、多くの反応中間体と思われる構造が得られた。具体的にはリブロース1,5-ビスリン酸と構造の良く似たカルボキシアラビニトールビスリン酸 (CABP) やマグネシウム-二酸化炭素複合体、カルシウム-二酸化炭素複合体、キシルロースビスリン酸 (XuBP)、そして生成産物である3-ホスホグリセリン酸 (PGA) などである。
また、2001年に古細菌型である Thermococcus kodakaraensis のForm III RubisCOの立体構造が明らかになった。遺伝子配列など、既知のRubisCOと相同性の低かったForm IIIは立体構造も他のFormと大きく異なっており、同一のサブユニット2個が対称性を持って結合しているL2単位が計5個対称性を持って結合している分子量497 kDa程度のL10型であることが明らかになった。
構造生物学的な触媒過程
編集

マグネシウムイオンを中心に多数の水素結合が存在している。また、リブロース1,5-ビスリン酸の2位および201番目のリシン残基にカルボキシル基(青色)が結合していることに注目。また、リブロース1,5-ビスリン酸の加水分解を受ける部位の近傍に379番目のセリン残基が存在し、加水分解活性に関わっているのではないかと考えられている。
立体構造が明らかになると同時に、サブユニットの機能についても明らかになった。Form Iにおける大サブユニットはカルボキシラーゼ/オキシゲナーゼ活性を有する触媒部位を有し、小サブユニットは機能未知な部分が多いが反応の調節を行なっているのではないかと考えられている。小サブユニットの無いForm I RubisCOでは活性が1%程度に落ち込むことが大腸菌発現系を用いた研究でわかっており、何らかの形でカルボキシラーゼ活性に関わっていることも示唆された。
Form II、IIIのサブユニットはForm I 大サブユニットと同様、触媒部位を有しており、基本的には以下に述べる基質の結合過程に従うと考えられる。触媒過程における最も多くの反応中間体が得られているのはホウレンソウのForm I RubisCOである。
- RubisCOへの基質結合に際して、まずK201(大サブユニットアミノ酸配列201番目のリシン)が二酸化炭素によってカルバミル化される。
- カルバミル化されたK201、D203(アスパラギン酸)、E204(グルタミン酸)によってマグネシウムイオンが水素結合により配位されRubisCOの活性型をとる。
- 2-カルボキシアラビニトール1,5-ビスリン酸(RuBPと構造の良く似た糖;CABP)は以下の水素結合によりRubisCOと結合する。
- 1位炭素に配位した酸素とK175およびマグネシウムイオン
- 3位炭素に配位した酸素とカルバミル化したK201、H327(ヒスチジン)およびマグネシウムイオン
- 5位炭素に配位した酸素とH327
- 1位リン酸に配位した酸素とK334
- リブロース1,5-ビスリン酸の3-オキソ中間体の2位炭素のカルボキシル基(すなわち取り込まれた二酸化炭素)に該当するCABPの2位カルボキシル基はマグネシウムイオンと水素結合する。
- リシンへの二酸化炭素の配位、マグネシウムイオン、CABPが結合するとloop6と呼ばれる通常活性中心にかぶさるように存在している部位がコンフォメーション変化し、生成物を開放できるようにopenな構造になる。
- 2分子の3-ホスホグリセリン酸が配位しているケースでもloop6はopenな構造を取っている。
これらの反応素過程において注意すべきなのは、K201にカルバミル化される二酸化炭素は基質として取り込まれるわけではない、という点である。基質として取り込まれるべき二酸化炭素は、Mg2+-CO2複合体および糖が配位したときに、3-オキソ中間体となるようリブロース1,5-ビスリン酸の2位にカルボキシル化される。
遺伝子改変系による機能改良
編集RuBisCOは上述したように、リブロース1,5-ビスリン酸へのカルボキシラーゼ反応という特有の反応を触媒するが、同時にオキシゲナーゼ反応も担うという酵素的な欠点も抱えている。また、比活性も低くカルビン-ベンソン回路の律速段階となっている。RubisCOのこうした欠点を克服することは主要作物の増産や地球温暖化防止の点においても極めて大きな影響を与えると考えられ、大腸菌を用いた遺伝子の突然変異法などによりその機能の改良が試みられてきた。
近年のRubisCOの改良に関する総説によると[1]、求められるべきPerfect RubisCOとは以下の条件を満たすものである。
- CO2への高い比活性および低いミカエリス・メンテン定数を有すること(具体的な数字として kcat/Km = 108 M−1s−1)
- ただし、CO2のミカエリス-メンテン定数は葉緑体ストロマにおけるCO2よりも高い値 (>8 μM) を示すこと(ストロマ中におけるCO2濃度が一定に保たれ常に最大速度を示すため)
- O2に対するCO2の特異性が無限大に大きいこと(オキシゲナーゼ活性が発生しないように、Sc/o = ∞)
現実のRubisCOは紅色非硫黄細菌 Rhodospirillum rubrum の場合、比活性が高いもののミカエリス-メンテン定数が高く、高CO2濃度に適応している。一方、タバコのRubisCOは比活性は低いが、ミカエリス-メンテン定数が低いため、低CO2濃度に適応している。この結果、現状で最も1.の条件を満たしているRubisCOはタバコとなりその値は kcat/Km = 3.2 × 105 である。タバコRubisCOのミカエリス-メンテン定数 (Km = 10.7 μM) を参考にした場合、逆算されうるPerfect RubisCOの比活性は1070 s−1となり、既存のRubisCOの100倍以上である。
2006年にシアノバクテリア Synechococcus PCC7492のRubisCO遺伝子の大量のランダム変異体を獲得し、大腸菌内でRubisCOが機能する場合にのみペントースリン酸経路の一部を用いたカルビン-ベンソン回路によって発現宿主が生育可能になる系にて優れたRubisCOの選抜を行なった報告がある[2]。本系では、RubisCO遺伝子の変異のみで発現量が野生型の約15倍あるいは比活性が5倍になるRubisCO変異体が得られている。このような、発現量の増大や比活性の向上が見られる理由は構造生物学的に明らかになっていないが、RubisCO自身の安定性に寄与している変異が導入されている事が一因と考えられている。本系を上手く応用することにより、今後さらに優れたRubisCO変異体が得られるのではないかと期待されている。
参考文献
編集- ^ Andrews, T. J.; Whitney, S. M. (2003). "Manipulating ribulose bisphosphate carboxylase/oxygenase in the chloroplasts of higher plants." Arch. Biochem. Biophys. 414: 159–169. PMID 12781767.
- ^ Parikh, M. R.; Greene, D. N.; Woods, K. K.; Matsumura, I. (2006). "Directed evolution of RuBisCO hypermorphs through genetic selection in engineered E.coli." Protein Eng. Des. Sel. 19: 113-119. PMID 16423843.