脊髄
脊髄(せきずい、英: spinal cord)とは、脊椎動物の脊椎の内側にある脊椎管の中に持つ、髄膜の内側に存在するニューロンと神経線維の集合体である。脳と脊髄とを併せて、中枢神経系と称する。脊髄は脊椎管の中から脊髄神経の枝を何本も出し、末梢と脳との橋渡しをする。ただし、脳と末梢とを繋ぐ神経は脊髄だけではなく、脳神経と呼ばれる脳から直接出ている神経も存在する。なお、脊髄は反射の一部も担っている。

構造
編集ヒトの脊髄は、延髄の尾側に始まり、第一腰椎と第二腰椎の間の高位で脊髄円錐となって終わり、終糸と呼ばれるひも状の繊維につながっている。
脊髄から直接出ている神経は神経根と呼ばれ、神経根が脊髄腔から出る高位によって、頸髄、胸髄、腰髄、仙髄、尾髄に分けられる。ただし人間では、尾髄は退化的である。脊髄は脊椎より短いため、脊髄の高位と脊椎のそれとは一致しておらず、脊髄の末端より下位の脊髄腔には神経根のみが伸びており、馬尾と呼ばれる。
脊髄の断面
編集

脊髄の断面は、縦走する神経細胞で構成される白質が、神経核(神経細胞体の集まり)である灰白質を囲む構造となっている。なお、逆に脳では灰白質が白質を囲む構造となっている。中央には第4脳室の続きである中心管がみられ、脳脊髄液で満たされる。灰白質部分は、(図6下側より)前角、側角、後角に分かれ、脳から降りてきた運動に関わる神経は脊髄前角で運動ニューロンにシナプスを作って連絡する。この連絡箇所は上肢・下肢に向かうニューロンが出る際に多くなり、頚部には頚膨大、腰部には腰膨大の膨らみとして肉眼でも観察できる。
灰白質は、存在する神経細胞の性質によって分類される。前角には遠心性神経の細胞体があり、脳から降りてきた運動に関わる神経は脊髄前角で下位運動ニューロンにシナプスを作って連絡する。後角には末梢から入る求心性神経とシナプスを形成する神経細胞体がある。また腰髄・胸髄だけに側角があり、ここには交感神経の神経細胞体がある。灰白質はさらに組織学的に幾つかの部分に分類される。さまざまな分類方法があるが、最も広く用いられているのは、ブロール・レクセドによる層分類である。レクセドはネコの脊髄を使って灰白質をIからXまでの10層に分類した[1][2][3]。
白質は大きく前索、側索、後索に分けられる。それぞれ、前索は前根の間、側索は前根と後根の間、後索は後根の間に概ね相当する。前索には上行路として前脊髄視床路、下行路として内側縦束、前皮質脊髄路、視蓋脊髄路、橋網様体脊髄路、前庭脊髄路、延髄網様体脊髄路がある。側索には上行路として脊髄視蓋路、脊髄オリーブ路、外側脊髄視床路、前脊髄小脳路、後脊髄小脳路が、下行路として赤核脊髄路、外側皮質脊髄路がある。後索には上行路として薄束、楔状束が、下行路として半円束、中間縁束がある。また、前索、側索、後索のいずれの部分でも、灰白質と接した部位は脊髄内の上下の連絡を行う神経線維の通る部分で、ここを固有束という。(主な伝導路については後述する)
脊髄は髄膜と呼ばれる三層の膜に包まれており、外側から硬膜(脊髄硬膜)、クモ膜(脊髄クモ膜)、軟膜(脊髄軟膜)と呼ぶ。すべての髄膜は大脳半球および脳幹を包むそれぞれの膜と一体になっている。脳と同様、クモ膜下腔には脳脊髄液が存在する。神経根の間の軟膜の外側は歯状靱帯となって硬膜に付着し、脊髄を固定している。硬膜は第二仙髄の高位で閉じている。
脊髄の分節
編集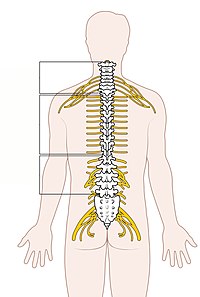
-
頸髄胸髄腰髄仙髄
- 画像ファイル(環境により文字がずれることもあります)

ヒトの脊髄は31の分節に分かれており(これを髄節と呼ぶ)、それぞれの髄節の左右の腹側から運動神経根が、背側から感覚神経根が末梢に出ている。腹側神経根と背側神経根はやがて合わさって脊髄神経となる。
31対の脊髄髄節はヒトでは以下に分類される。
- 8対の頸髄(第一頸神経は後頭骨と第一頸椎(環椎)の間から、第二頸神経以下はそれぞれ一つ上の高位の頸椎の下(椎間孔)から出る)
- 12対の胸髄(第一〜第十二胸神経は、それぞれ第一胸椎〜第十二胸椎の下から出る)
- 5対の腰髄(第一〜第五腰神経は、それぞれ第一腰椎〜第五腰椎の下から出る)
- 5対の仙髄(第一〜第五仙骨神経は、それぞれ第一仙椎〜第五仙椎の下から出る)
- 1対の尾髄(尾骨神経は尾椎から出る)
- ただし、第一頸髄と尾髄では、背側神経根はなく運動神経根だけが出る。
脊椎は脊髄に比べて成長が早く、最終的に長くなるため、成人では下位の脊髄髄節になるほど、対応する高位の脊椎骨に比べて高い位置にある。発生のはじめ(胎生3ヶ月まで)では脊椎と脊髄の高さは一致しているが、成人の脊髄の終わり(脊髄円錐)はだいたい第一腰椎と第二腰椎の間になる。腰仙髄は第九胸椎から第二腰椎の間にあり、例えば第四仙髄の神経根は胸腰椎移行部のあたりで脊髄から出て脊柱管内を下行し、第四仙椎の下から出る。脊髄円錐より下は馬尾と呼ばれる。
脊髄の径は、次の二つの部位で大きくなっている。
- 頸膨大 - 大体腕神経叢を構成する神経が出る髄節に一致し、上肢を支配する。第四頸髄から第一胸髄であり、椎骨の高さもほぼ一致している。
- 腰膨大 - 腰仙骨神経叢(腰神経叢、仙骨神経叢)を構成する神経が出る髄節に一致し、第二腰髄から第三仙髄にあたる。脊椎高位は第九から第十二胸椎の高さである。

発生
編集脊髄は神経管の一部から作られる。神経管が形成され始めると、脊索はソニック・ヘッジホッグ (SHH) を分泌する。これによって底板(神経管の腹側正中にできる部分)が誘導され、さらに底板からもSHHが分泌されて基板(神経管腹側の主に運動神経となるもとの部分)が誘導される。一方外胚葉からは骨形成タンパク質 (BMP) が分泌され、これは蓋板(背側正中にあり、翼板を左右に分ける)を誘導し、さらに蓋板もBMPを分泌して翼板(神経管背側の主に感覚神経となるもとの部分)が誘導される。翼板と基板は境界溝で区切られている。
また底板はネトリンタンパク質も分泌する。ネトリンは翼板の神経細胞のうち、温痛覚ニューロン(後述)に正の走化性を起こす誘引物質として働き、この神経の軸索を中心管の前(脊髄白前交連という)で交叉して反対側の腹側を上行し、視床まで進ませる(前脊髄視床路という、後述)働きをする。
神経系が正しく組み立てられるには、神経細胞でもプログラム細胞死 (programmed cell death, PCD) によって細胞が削ぎ落とされる必要があり、これはかつてヴィクトル・ハンブルガーとリータ・レーヴィ=モンタルチーニによってニワトリで研究されたが、最近の研究でも証明されている。

感覚系伝導路
編集体性感覚性神経系には、触覚・固有覚(位置覚と振動覚)を伝える後索・内側毛帯路と、温痛覚(温度覚と痛覚)を伝える脊髄視床路の二つの伝導路がある。
どちらの伝導路も、感覚受容器から大脳皮質に情報が伝わるまでに3種類のニューロンがかかわっている。このニューロンを末梢から順に一次・二次・三次ニューロンという。一次ニューロンは脊髄後根神経節に細胞体があり、そこから末梢の感覚受容器と脊髄の両側に軸索を伸ばしている。
触覚・固有覚の経路(後索・内側毛帯路)では、一次ニューロンは脊髄に入ると同側の後索を上行する。第六胸髄以下のニューロンは後索の内側寄りにある薄束を通る。それより上のニューロンは後索の外側にある楔状束を通る。一次ニューロンは延髄に入るとそれぞれ薄束核、楔状束核と呼ばれる神経核でシナプスを形成しニューロンを交代する。二次ニューロンは延髄で交叉(左右のニューロンが入れ替わる)し、内側毛帯と呼ばれる束になってさらに上行して視床の後外側腹側核(VPL核)に入り、三次ニューロンとシナプスを形成する。三次ニューロンは視床から大脳の内包後脚を通り、頭頂葉の中心後回(ブロードマンの脳地図の3,1,2野)にある感覚野に至る。
温痛覚伝導路は、また異なった経路を通る。痛覚の一次ニューロンは脊髄に入ると1-2髄節上行または下行して後角膠様質でシナプスを形成する(この数髄節上行または下行する経路を後外側路あるいはリサウエル路という)。シナプス後の二次ニューロンは対側に交叉し、脊髄の前外側を視床まで上行する(前脊髄視床路が触覚を、外側脊髄視床路が温痛覚を伝える)。このため、脊髄視床路は前外側系とも呼ばれる。二次ニューロンは視床の後外側腹側核(VPL核)で三次ニューロンとシナプスを形成する。三次ニューロンは内包後脚を通って、頭頂葉の中心後回(ブロードマンの脳地図の3,1,2野)にある感覚野に投射する。
前外側系のニューロンは視床に投射するだけではなく、途中で脳幹網様体にも軸索が伸びている(脊髄網様体路)。この網様体からはさらに海馬(痛みの記憶を形成する)、視床の正中中心核(CM核、漠然とした痛みを感じる)など大脳や間脳のさまざまな場所に投射している。中脳中心灰白質(中脳水道周囲灰白質)にも二次ニューロンは投射しており、ここから延髄の大縫線核、さらに脊髄へと投射があって、痛覚信号の入力があるとそれを抑制する働き(負のフィードバック)を持つ。これによって痛み刺激が軽減される。
運動系伝導路
編集
上位運動ニューロンによる入力は、大脳皮質および脳幹の神経核から来る。軟質骨上位運動ニューロンは骨髄脊髄路と呼ばれる。ブロードマンの脳地図の4野(一次運動野)、6野(運動前野)、1-3野(体性感覚野)および5野(頭頂葉体性感覚連合野)に始まり[4][5]、内包の膝および後脚を通って大脳脚、橋から延髄の錐体に至る。ここで運動ニューロンの軸索のうち約85%が対側に交叉し、脊髄の外側皮質脊髄路を下行する。残りの約15%のニューロンは同側の前皮質脊髄路を下行する。その名と異なって、中心前回(一次運動野)第V層のベッツ細胞(巨大錐体細胞)に始まる上位運動ニューロンは数の上では少数であり、ヒト以外の霊長類では60%以上の上位運動ニューロンが運動前野から起始していることが明らかになっている[6]。
脳幹からは4つの運動伝導路が上位運動ニューロンを出している。赤核脊髄路(中脳の赤核より)、前庭脊髄路(橋延髄境界にある外側前庭核より)、視蓋脊髄路(中脳の上丘より)、網様体脊髄路(橋および延髄の網様体からそれぞれ)の4つである。赤核脊髄路は側索(皮質脊髄路のすぐ前方)を、その他の運動路は前索を下行する。
下位運動ニューロンの機能は、外側皮質脊髄路および前皮質脊髄路の二つで異なっている。外側皮質脊髄路の上位運動ニューロンの軸索は、脊髄前角のうち背外側 (DL) 下位運動ニューロンとシナプスを形成する。DLニューロンは四肢遠位の巧緻運動を支配する。四肢に関係するので、DLニューロンは脊髄のうち頸膨大および腰膨大の灰白質にのみ存在する。外側皮質脊髄路は延髄錐体で交叉した後は、脊髄では交叉しない。
一方前皮質脊髄路の上位運動ニューロンは、大脳皮質と同側の脊髄前索を下行し、その軸索は同側の脊髄前角のうち腹内側 (VM) 下位運動ニューロンとシナプスを形成するか、あるいは白前交連で対側に交叉した後、その高位の髄節内にあるVM下位運動ニューロンとシナプスを形成する。脳幹に始まる赤核脊髄路、視蓋脊髄路および前庭脊髄路の上位運動ニューロンも同側の脊髄前索を下行するが、これらのニューロンは交叉を行わず、同側の脊髄前角にあるVMニューロンとシナプスを形成する。VM下位運動ニューロンは、体幹の大きな姿勢筋(脊柱起立筋など)を支配する。VM下位運動ニューロンはDLニューロンと異なり、脊髄のすべての高位の前角内に見られる。
脊髄小脳路
編集固有覚の情報は、大脳皮質だけでなく小脳へも伝えられ、その経路は3つある。第二腰髄以下の固有覚情報は、感覚受容器(ゴルジ腱器官や筋紡錘)から腹側脊髄小脳路または前脊髄小脳路と呼ばれる経路を通って脊髄に入る。一次ニューロンの神経細胞体は脊髄後根神経節にある。脊髄に入ると、二次ニューロンとシナプスを形成する。二次ニューロンの神経細胞体は脊髄前角の辺縁にあり、脊髄境界細胞と呼ばれる[7]。二次ニューロンは交叉して対側にわたり、側索の前外側辺縁を上行し、橋上部の上小脳脚から小脳に入る。多くの神経線維は再び交叉して小脳虫部前部に投射する。
第二腰髄から第一胸髄までの固有覚を伝える一次ニューロンは、脊髄後角基部の背核(胸髄核、クラーク核とも)とシナプスを形成する。二次ニューロンは交叉せず、同側の脊髄側索の後外側を上行し、下小脳脚から小脳に入って小脳虫部の吻側部および尾側部に投射する。この経路は背側脊髄小脳路(後側脊髄小脳路)と呼ばれる。
第八頸髄から上位の固有覚一次ニューロンは、脊髄に入るとそのまま後索を上行し、延髄の副楔状束核(外側楔状束核)でシナプスを形成する。二次ニューロンは下小脳脚から小脳に入り、同側の小脳半球皮質の
らかの理由による切断で起こる。椎骨や椎間板が破壊され、骨の鋭利な破片が脊髄を傷害することもある。脊髄損傷を被った人は、通常体の一部の感覚低下がある。さらに軽症で手や足の機能の障害、重症の場合は対麻痺や四肢麻痺、あるいは損傷部位以下の全機能の喪失が起き、最悪の場合呼吸困難を引き起こし死に至る場合もある。
上位運動ニューロン線維の損傷があると、傷害部位と同側に特徴的な症状が現れる。腱反射の亢進や筋緊張の亢進、あるいは筋力の低下などが起きる。また下位運動ニューロン線維の損傷の場合にも特徴的な症状が出る。障害側の全体ではなく、傷害を受けた神経の支配筋だけが影響を受ける。また筋力低下、筋緊張の低下、腱反射の減弱や消失、筋萎縮などが起きる。
脊髄損傷をもっとも起こしやすい部位は、頸髄および腰髄である。
追加画像
編集-
 図7 脊髄の模式図。
図7 脊髄の模式図。 -
 図8 各レベルの脊髄横断面。
図8 各レベルの脊髄横断面。
脚注
編集- ^ Rexed B. The cytoarchitectonic organization of the spinal cord in the cat. J Comp Neurol 1952;96:415-495.
- ^ Rexed B. A cytoarchitectonic atlas of the spinal cord inth cat. J Comp Neurol 1954;100:297-379.
- ^ Rexed B. Some aspects of the cytoarchitectonics and synaptology of the spinal cord. In: Eccles JC, Shadé JP, eds. Progress in brain research. Vol. II. Organization of the spinal cord. Amsterdam: Elsevier, 1964:58-92.
- ^ Coulder JD, Ewing L, Carter C. Origin of primary sensorimotor cortical projection to lumbar spinal cord of cat and monkey. Brain Res 1976;103:366-372
- ^ Jones EG, Wise SP. Size, laminar and columnar distribution of efferent cells in the sensory-motor cortex of monkeys. J Comp Neurol 1977;175:391-438
- ^ Dum RP, Strick PL. The origin of corticospinal projections from the premotor areas in the frontal lobe. J Neurosci 1991;11:667-689
- ^ Cooper S, Sherrington CS. Gower's tract and spinal border cells. Brain 1940;63:123-134.