ディコトモクラディウム
ディコトモクラディウム Dichotomocladium は、ケカビ目の菌類の1つ。胞子形成部の枝が二又分枝し、その多くの先端が不実の棘となる。無性生殖は単胞子性の小胞子嚢を小さな頂嚢の表面に多数つける。
| ディコトモクラディウム属 | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
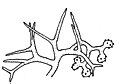
 Dichotomocladium elegans の胞子形成部
| ||||||||||||||||||
| 分類(目以上はHibbett et al. 2007) | ||||||||||||||||||
| ||||||||||||||||||
| 学名 | ||||||||||||||||||
| Dichotomocladium Benny & Benjamin 1975 | ||||||||||||||||||
| 種 | ||||||||||||||||||
|
下記参照 |
具体的な形質
編集タイプ種である D. elegans を元に記す[1]。
菌糸体
編集
通常の寒天培地上で培養は可能だがその生育は限定的で、合成ムコール培地(ケカビ類の培養に標準的に用いられる培地)で26℃、6日間の培養でもコロニーの径は3cm程にしかならない。房状で密集したコロニーを形成し、当初は白く、後にオリーブ色っぽく色づいてくる。
無性生殖器官
編集無性生殖は単胞子性の小胞子嚢による。胞子形成柄は直立するか、斜めに立ち上がり、高さ1-2cmに達し、その径は5-8㎛ほど。透明から明るい黄色を呈し、当初は無隔壁だが後に隔壁を不規則に生じる。柄は単一か、あるいは不規則に仮軸状の分岐を見せる。細胞壁は厚くて表面に細かな凹凸がある。この柄の上に胞子形成部分を2-18個ほどつける。この部分は柄の上にある程度一定の間隔を置いて単独に、あるいは対をなし、あるいは輪生状に生じる。それらは横枝として出た後に短い間隔で二叉分枝を繰り返し、25-45×4-8.5㎛の横枝が混み合って胞子形成部を作り上げる。胞子形成部は成熟時には灰色から茶褐色になり、その概形は不規則な塊状からやや球形になり、その径は160-250㎛に達する。それを構成する菌糸は分枝を繰り返し、その枝の多くは先端が伸張して不実の棘状突起となり、先端近くの数個の横枝だけが胞子形成枝となる。不実の棘は長さ6-32㎛、基部の太さが1-3㎛。
胞子形成部の枝の最後から二番目の分枝には普通は1つの隔壁があり、最後の分枝は径が1-3×3㎛で、その先端が膨らんで球形からやや角張った膨らみとなって表面に突起を並べた頂嚢となり、その径は4-6㎛。表面には柄でつながった小胞子嚢が6-8個ほど生じる。小胞子嚢の柄は長さ1-2㎛で先端に向かって細まっている。小胞子嚢は卵形から楕円形で長さ5-6㎛、幅3.5-4.5㎛で無色からやや黄色く色づく。表面の壁は薄くて小さな棘状突起が並んでいる。柱軸は凹んでおり、径2㎛。胞子嚢胞子は小胞子嚢と大きさ、形共にほぼ同じで、表面の壁は薄くて滑らかとなっている。
-
 胞子形成部の枝の一部
胞子形成部の枝の一部 -
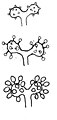 頂嚢と小胞子嚢の発達・前半
頂嚢と小胞子嚢の発達・前半 -
 同・後半
同・後半



有性生殖
編集有性生殖は接合胞子嚢の形成による。自家不和合性なので好適な株同士を合わせたときのみ接合は行われる。接合胞子嚢は球形から亜球形で径40-60㎛、表面の壁は茶色でやや透明、表面には高さ2-5㎛の突起が一面にある。その外側に残る配偶子嚢は茶色をしていて長さ2-5㎛、支持柄は対面型、いわゆるH字型になる。
特徴まとめ
編集新属の記載文としては以下のようになっている[2]。
胞子形成菌糸は基質菌糸から直接に出てまっすぐか斜めに立ち上がり、単一か分枝し、胞子形成部を側面から出すか、あるいは先端の上に出す。胞子形成部は数回二叉分枝をした構造からなり、その枝の一部は伸び出て不実の棘状突起となり、そうでないものはその先端が膨らんで角張った、あるいは丸っこい頂嚢を形成し、この頂嚢の上に小さな柄のある小胞子嚢が形成される。不実の棘状突起は真っ直ぐか曲がりを持ち、表面に配置する。小胞子嚢の柄は細くて円筒形で、先細りとなっている。小胞子嚢は単胞子性でほんの小さな柱軸があり、球形から楕円形で、表面の壁は細かな棘状突起があり、薄くて持続性である[3]。柱軸は切れ込みがあって凹んでいる。胞子嚢胞子は大きさ、形共に小胞子嚢とほぼ同じ。接合胞子嚢は球形から亜球形で壁は暗色、粗い突起によって表面が装飾されている。配偶子嚢は明かな形で残り、暗色で表面は滑らかかざらつく。接合子はH字型。
学名の意味は dichotomus (2つに割れた)と cladodes(枝)を合わせたもので、胞子形成部の側枝が二叉分枝をしていることに基づく。
分布と生態
編集タイプ種のホロタイプは北アメリカ、カリフォルニアのネズミの糞から分離されたもので、同時に調べられた株は北アメリカからメキシコに渡る諸地域で、やはりネズミなどの糞から多くが発見されている[4]。同時に記載されたもう1種、 D. robustum もやはりメキシコから北アメリカに渡ってのネズミ等の糞から発見されている[5]。また別属から本属へ移動された D. hesseltinii はインドからのみ知られ、土壌とウサギの糞から取り出されたものであった[6]。後に追加して記載された2種もそれぞれ動物の糞から発見されており[7]、総じて温暖な地域の動物の糞に出現する糞生菌とみられる。
下位分類
編集この属が記載されたとき、その下に置かれたのは以下の3種であった[8]。
- Dichotomocladium
- D. elegans
- D. hesseltinei
- D. robustum
これらは胞子形成部の特徴や小胞子嚢をつける頂嚢の形などで識別され、D. hesseltinei は胞子形成部のまとまりがゆるくて分枝が見て取りやすく、棘状突起が少ないことや頂嚢が角張っていることで、また D. robustum は胞子形成部が大きくて、しかし胞子形成柄に対して少数しかつけないことなどで区別される。
後に同一の著者らによって以下の2種が追加された[7]。
- D. floridanum
- D. sphaerosporum
D. floridanum は北アメリカで糞から得られたもので D. robstum に似ているが小胞子嚢が卵形をしている。D. sphaerosporum は D. elegans に似ているが小胞子嚢が球形をしているものでパキスタンの糞から得られた。
類似の群
編集本属に特によく似たものにイトエダカビ属 Chaetocladium がある[9]。本属とは以下の点で共通の特徴を持っている。
- 無性生殖が単胞子性の小胞子嚢のみであり、大きな胞子嚢は作らないこと。
- 小胞子嚢を小さな頂嚢の表面に多数作ること。
- 胞子形成柄に側枝が出て、その側枝が繰り返し分枝し、その先端に一部は棘状突起、残りは小胞子嚢をつける頂嚢ができること。
このような点から両者はよく似ており、実際に現在は本属のものとされている D. hesseltinei は新種記載の時点ではこの属のものと判断されて C. hesseltinei と命名されていた。
しかし本属はこの属のものとは以下の点で異なっている。
- 胞子形成部の側枝の分枝の様子が異なる。
- イトエダカビ属では側枝は輪生状に出る。まず胞子形成柄のある位置から2本ないしそれ以上の数の枝が菌糸の周囲から外向けに伸び、またその先端より下の位置から側枝を輪生状に出し、これを数度繰り返し、最後にそれらの側枝の先端に多少膨らんだ頂嚢が生じて小胞子嚢を形成し、最後の側枝の先端は棘状突起になる。
- 本属では側枝は二叉分枝をする。側枝は10-15回、時にはそれ以上にわたって二又に分枝を繰り返し、次第に全体としてカーブを描き、その先端の方に小胞子嚢をつける頂嚢ができる。それによって胞子を形成する部分は内側に、棘状突起は外側に向かい、全体としては球状の胞子形成部になる。
- 接合胞子嚢についても若干の違いがある。
- 形態以外の性質として、イトエダカビ属のものは低温を好み、7℃でよく生育し、20℃を超えると著しく生育が阻害されるが、本属の種は7℃では生育がよくなく、18℃から30℃までの範囲でとてもよい発育を示す。
- イトエダカビ属のものは他のケカビ類を宿主とする条件的寄生菌[10]で、宿主の上に虫瘤状の構造を作ることが知られるが、本属のものは寄生性を示さない。
分類
編集本属が記載された時点では、本属はさほどの議論抜きでエダケカビ科 Thamnidiaceae に所属させられた。これは主として無性生殖器官の構造に基づくもので、それ以前より小胞子嚢を形成するケカビ類はエダケカビ科とすることが行われてきた。本属を記載したBenny & Benjamin(1975)の定義では『大型の胞子嚢(ないこともある)と少数胞子、あるいは単胞子の小胞子嚢を形成し、小胞子嚢は単胞子の場合でも胞子嚢壁と胞子が区別できる。胞子嚢は顕微鏡下で滑らか。配偶子嚢壁は有色で厚壁で表面は突起に覆われる。それを支える配偶子嚢は両方同型で付属突起を持たず、接合はH字型』というものである。これは小胞子嚢を形成するケカビ類からコウガイケカビ科やクスダマカビ、ラジオミセス属などを排除するような形である。
イトエダカビはこの時点ではエダケカビ科に含めていたが、その後この属だけで単独にイトエダカビ科 Chaetocladiaceae が立てられると、Benny & Benjamin(1993)は本属に新種を追加すると同時に本属をこの科に含めるべきとの判断を示した。
しかし21世紀に入って発展した分子系統の研究により、このような分類体系が類縁関係を反映していない例が多いことが判明し、体系の見直しが行われた。Hoffmann et al.(2013)ではケカビ目全体にわたっての系統を分析して科の構成を検討しているが、本属については5種を使ってこれらが単系統をなすことが示された。同時に本属ともっとも近縁とされたのが Lichtheimia であった。これは元来はユミケカビ属 Absidia に含められていたものであり、その特徴は匍匐菌糸を出すこと、および大きな胞子嚢のみを形成し、胞子嚢には円錐形のアポフィシスがあって胞子嚢全体が洋なし型に近い形をしていることである。Lichtheimia に移された種は、この属の中でも性質が異なるとみられていたものの一部で、特に高温で生育が可能で、そのためもあって人間にケカビ症を引き起こすことが知られたものを含んでいる。したがって本属とは形態的には共通する点がごく少ないのであるが、分子系統の検討が進むにつれて両者の近縁性が指摘されてきた。それがあらためて確かめられた形である。両者には形態上の共通点はごく少ないが、共通な性質として高温への耐性があり、本属のものでも普通に35℃でよく成長し、一部では37℃でも成長が可能であることが確かめられている。この両者で1つの科 Lichthemaniaceae を立てることが提案され、またその中でそれぞれの属を別個の亜科とし、本属は Dichotomocladioideae の下に置くこともまた提起されている。
ちなみにこの Hoffmann et al.(2013) の結果によると、イトエダカビ属はケカビ属を含むケカビ目の多くのものが集中するクレードに位置しており、本属の属するクレードとはケカビ目の系統樹の基底近くで分岐したことになっている。
出典
編集- ^ 以下、Benny & Benjamin(1975),p.340
- ^ Benny & Benjamin(1975),p.338
- ^ 壁が割れたり溶けたりして胞子を放出するようなことをしない、という意味である。
- ^ Benny & Benjamin(1975),p.340-342
- ^ Benny & Benjamin(1975),p.343
- ^ Benny & Benjamin(1975),p.347
- ^ a b Benny & Benjamin(1993)
- ^ Benny & Benjamin(1975),p.339
- ^ 以下、Benny & Benjamin(1975),p.338-339
- ^ 寄生性を示すが、培地上で単独に生育させることもできる。
参考文献
編集- Gerald L. Benny & R. K. Benjamin, 1975. Observations on Thamnidiaceae (mucorales), New Taxa, New Combinations, and notes on selected species. Aliso Vol. 8, No. 3, pp.301-351.
- Gerald L. Benny & R. K. Benjamin, 1975. Observations on Thamnidiaceae (mucorales), VI. Two new species of Dichotomocladium and The Zygospores of D. hesseltinei(Chaetocladiaceae). Mycologia Vol.85 issue4 p.660-671.
- K. Hoffmann et al. 2013. The family structure of the Mucorales: a synoptic revision based on comprehensive multigene-genealogies. Persoonia 30:p.57-76.